2019年4月10日,, 北京中醫(yī)藥大學(xué)徐安龍教授團(tuán)隊(duì)與耶魯大學(xué)David G. Schatz教授和熊勇教授團(tuán)隊(duì)合作在抗體和T細(xì)胞受體V(D)J重排機(jī)制演化研究領(lǐng)域再獲新進(jìn)展,研究成果以長文形式(Article)在國際權(quán)威學(xué)術(shù)期刊Nature在線發(fā)表,,論文題目為“Transposon Molecular Domestication and the Evolution of the RAG Recombinase”,,該成果揭示了原始轉(zhuǎn)座酶protoRAG演化為重組酶RAG的重要機(jī)制。

免疫系統(tǒng)包括天然(非特異性)免疫系統(tǒng)和適應(yīng)性(特異性)免疫系統(tǒng),。人類適應(yīng)性免疫的關(guān)鍵機(jī)制是抗體和T細(xì)胞受體重排激活基因 (Recombination activating gene,, RAG)介導(dǎo)的V(D)J重排機(jī)制。該機(jī)制是由美籍日裔科學(xué)家Susumu Tonegawa (利根川進(jìn))團(tuán)隊(duì)在1979年發(fā)現(xiàn),, 他因此于1987年獲得諾貝爾生理學(xué)或醫(yī)學(xué)獎,。 同時(shí), 利根川進(jìn)創(chuàng)造性地提出了重排機(jī)制的轉(zhuǎn)座子起源假說。數(shù)十年來,,一代又一代學(xué)者不斷求索,,探求重排機(jī)制的起源。 直到2016年,, 徐安龍教授領(lǐng)導(dǎo)的中山大學(xué)團(tuán)隊(duì)在文昌魚的基因組中發(fā)現(xiàn)了六億年前RAG轉(zhuǎn)座子的“分子活化石”——ProtoRAG,, 為轉(zhuǎn)座子起源假說提供了直接的證據(jù), 文章發(fā)表在Cell上, 并被同期Cell雜志在Leading edge專欄點(diǎn)評。
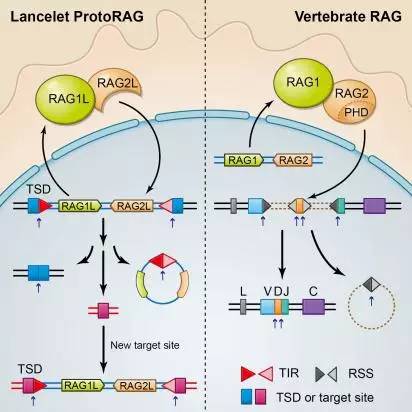
文昌魚ProtoRAG轉(zhuǎn)座子和脊椎動物RAG蛋白的功能比較
(圖片引自徐安龍教授團(tuán)隊(duì)2016年Cell文章:Huang et.al, Cell, 2016.)
但是,, RAG究竟是怎樣在漫長的進(jìn)化中失去了轉(zhuǎn)座功能而只精確地履行免疫球蛋白(包括抗體和T細(xì)胞受體)基因重排功能的呢,?
為了回答這一問題,北京中醫(yī)藥大學(xué)與耶魯大學(xué)合作研究團(tuán)隊(duì)首先解析了文昌魚的ProtoRAG的冷凍電子顯微鏡結(jié)構(gòu),,通過與RAG結(jié)構(gòu)進(jìn)行比較鑒定出了在RAG中具有重要功能的氨基酸殘基和結(jié)構(gòu)域,。它們的存在、改變或丟失能夠改變RAG切割底物時(shí)具有的配對切割(Couple Cleavage)傾向和對非對稱底物的偏好,,甚至使RAG在細(xì)胞內(nèi)產(chǎn)生轉(zhuǎn)座活性,。尤其是RAG1中的第848位氨基酸——精氨酸和RAG2中的酸性氨基酸富集的結(jié)構(gòu)域(Acidic Hinge),二者共同作用使RAG在細(xì)胞中的轉(zhuǎn)座活性降低1000多倍,。最終,,合作研究團(tuán)隊(duì)提出了RAG家族為適應(yīng)不斷變化的環(huán)境和宿主需求持續(xù)演變的進(jìn)化模型。該研究成果可能為文昌魚的ProtoRAG演化到包括人在內(nèi)的有顎脊柱動物RAG提供了演化路徑,。
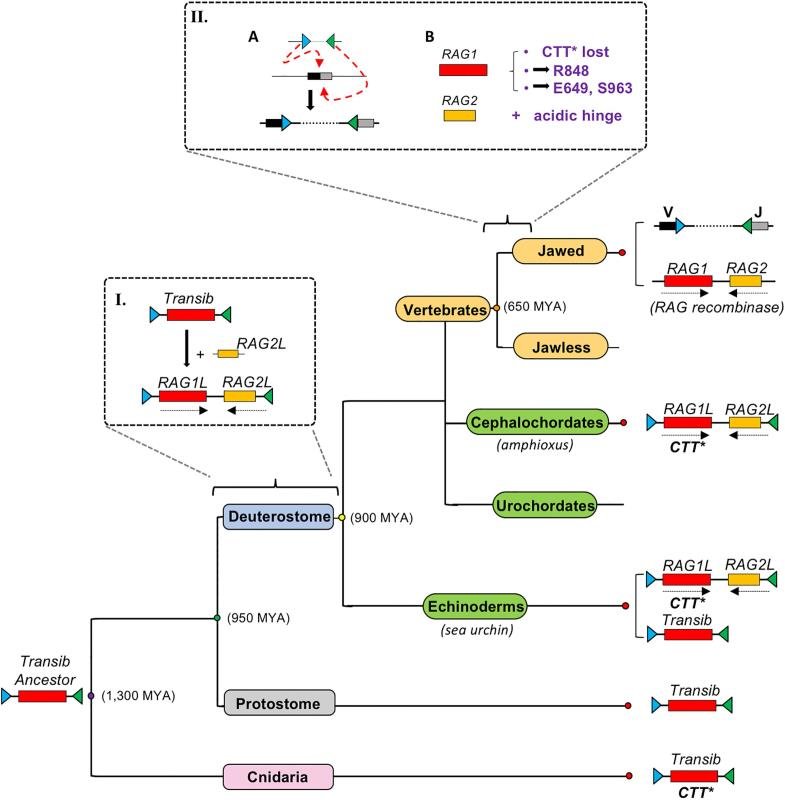
RAG進(jìn)化模型(圖片引自2019 Nature文章:Zhang et.al, Nature, 2019)
耶魯大學(xué)中國學(xué)者張宇航博士和Tat Cheung Cheng博士(中國香港)為論文的共同第一作者,, 北京中醫(yī)藥大學(xué)黃光瑞博士和陸清怡博士研究生為第三和第四作者; 北京中醫(yī)藥大學(xué)徐安龍教授,、耶魯大學(xué)David G. Schatz教授和熊勇教授為論文的共同通訊作者。

全文鏈接:https://www.nature.com/articles/s41586-019-1093-7